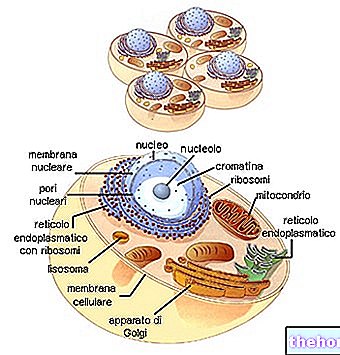
АСОЦІАЦІЯ (АБО "ЗВ'ЯЗАННЯ" АБО "ЗВ'ЯЗОК")
До сих пір ми обробляли схрещування дингібрида (або полігібрида) Менделя за припущенням, що різні алельні пари насправді зустрічаються на різних парах гомологічних хромосом. Але кількість хромосомних пар, хоча і відрізняється від виду до виду, змінюється у вузьких межах (кілька видів досягають близько ста хромосом), тоді як кількість генів можна порахувати десятками тисяч.
Те, що персонажі, обрані Менделем для своїх експериментів, відокремили всіх незалежно (без того, щоб заплутати розрахунки у розподілі фенотипів у F2 полігібридного схрещування), був щасливим шансом. Якщо дві пари алелей були знайдені на сусідніх локусах, закон наступне буде називатися законом асоціації.
Знаючи, що дуже багато персонажів мають своє місце розташування в одній хромосомі і що вони є хромосомними парами, які незалежно відокремлюються під час мейозу, можна добре зрозуміти, як часто трапляється, що дві пари символів, якщо вони були пов'язані в хромосомі з батьками так само в гаметі, а отже, і в організмі, до якого вона принесе свій власний генетичний матеріал.
Таким чином, ми бачимо, що "асоціація являє собою" виняток, далеко не рідкість, щодо незалежності, проголошеної в третьому законі Менделя.
ОБМІН АБО "ПЕРЕКРИСТАННЯ" ТА РЕКОМБІНАЦІЯ
Говорячи про мейоз, ми вказали, що існують два різні моменти змішування генетичного матеріалу: один - це поділ хромосом у гаметах, і той, який спостерігав Мендель.
Інший момент, який фактично передує, - це той, у якому чотири хроматиди кожної пари гомологічних хромосом взаємно обмінюються ідентичними ознаками. Після цього обміну два фактори, які були пов’язані в одній хромосомі, натомість будуть незалежними в гаметах. те, що відбувається обмін, є пропорційним, у першому наближенні, довжині хромосоми, а в більш довгих хромосомах може бути навіть більше, ніж обмін.
Це явище можна виявити цитологічно, спостерігаючи за достатньою кількістю мейозів під мікроскопом.
Частота повторного збігу-це швидкість, з якою будь-які два символи, які були пов’язані в батьківському поколінні, по-різному поєднуються у F2.
Якщо два локуси абсолютно суміжні, ймовірність того, що хіазма розділить їх, буде практично нульовою. Швидкість рекомбінації складе: n ° рекомбінант. Якщо два локуси знаходяться на двох різних хромосомах, швидкість рекомбінації становитиме 0,5 (рівна ймовірність, що два символи, які були об’єднані в поколінні Р, випадково опиняться разом у F2). Тому швидкість рекомбінації може коливатися від 0,0 до 0,5. Для невеликих відстаней на хромосомі відстань і швидкість рекомбінації прямо пропорційні. Для більших відстаней існує ймовірність того, що між двома локусами відбудуться два обміни. Тепер буде очевидно, що два фактори, розділені двома біржами, знову пов'язані. зрозуміло, що на цьому етапі втрачається пропорційність між відстанню локусів та ймовірністю рекомбінації.
Локуси, знайдені на одній хромосомі, становлять «групи асоціацій». Дуже віддалені локуси можуть мати таку ймовірність відокремлення шляхом обміну, що вони поводяться як незалежні, але кожен з них буде пов'язаний з меншою швидкістю рекомбінації з проміжними локусами.
Коли швидкість рекомбінації між багатьма парами генів всередині асоціативної групи відома, можна починати побудову «генетичних карт». Маючи на увазі, що відстань між двома генами (a і b) виражається швидкістю рекомбінації і що відстань a від третього гена c може бути або сумою, або різницею щодо його відстані від b, це можливо реконструювати карту взаємних відстаней, яка буде генетичною картою в межах цієї групи асоціацій, тобто цієї хромосоми.
Тепер ми повинні загалом розглянути деякі концепції, що обмежують фенотипний прояв генотипових ознак.
Перш за все ми поговоримо про поняття пенетрантності та експресивності, а потім приділимо особливу увагу феноменам регуляції дії генів.
ПЕНТРЕНС
Пенетрантність гена означає його здатність проявлятися у фенотипі. Проникність вимірюється статистично шляхом підрахунку частоти фенотипів, які показують цей характер із 100 генотипів, які його містять. Ознака з проникністю 0,7 - це ознака, яка фенотипово зустрічається у 70% її генотипічної частоти.
ВИРАЖНІСТЬ
Експресивність - це кількісна оцінка ступеня фенотипного прояву.
РЕГУЛЮВАННЯ ГЕННОЇ ДІЇ
Клітини виробляють всі свої ферменти та білки з однаковою швидкістю і в той же час. Наприклад, клітини кишкової палички можуть отримувати енергію та атоми вуглецю з дисахариду лактози, оскільки вони здатні розщеплювати їх на глюкозу та галактозу завдяки ферменту бета-галактозидази. У нормальній кишковій паличці, яка може містити лактозу, є приблизно 3000 молекул бета-галактозидази, що дорівнює 3% білків цієї клітини; за відсутності лактози буде лише одна молекула бета-галактозидази на бактеріальну клітину. Галактозидаза буде синтезована з нових молекул мРНК, коли її можна буде використовувати. Мутантні штами E. coli, багаті на фермент, відомі навіть за відсутності лактози: ці мутанти перебувають у невигідному становищі порівняно з нормальними клітинами, оскільки вони змушені до непотрібного споживання енергії та матеріалів для виробництва ферменту, який залишиться без субстрату. Речовини, які спричиняють збільшення кількості ферменту, як, наприклад, з лактозою, називатимуться індукторами, а ферменти - індуктивними. Інші речовини, також специфічно викликають вироблення певних ферментів. Також у кишкової палички, наприклад, здатної будувати всі свої амінокислоти, що мають вуглець та амоній (NH3), присутність у культуральному середовищі певної амінокислоти (наприклад, гістидину) блокує вироблення всіх ферментів, пов’язаних із біосинтез самої амінокислоти: про ці ферменти буде сказано, що вони репресуються.У бактеріальних клітинах молекули мРНК зриваються незабаром після їх утворення, і тому контроль виробництва мРНК означає контроль ферментативного синтезу одночасно час.
ОПЕРОН
Щоб пояснити, як бактеріальна клітина здатна контролювати власне виробництво ферментів, Якоб і Моно сформулювали гіпотезу оперону; оперон утворений кількома генами, які функціонально пов'язані і вирівнюються без перерв уздовж ділянки ДНК.Оперон складається з трьох різних типів генів: промотора, де починається утворення мРНК; оператор, де здійснюється контроль; один або кілька структурних генів, які кодують ферменти або інші білки. інші ферменти, що беруть участь у метаболізмі лактози.Ці гени прилягають один до одного і транскрибуються один за одним по одній спіралі ДНК в одну молекулу мРНК. Вироблені таким чином молекули мРНК активні протягом дуже короткого часу, після чого вони руйнуються специфічними ферментами.
Активність оперону, у свою чергу, контролюється іншим геном, регулятором, який також може бути віддалений від оперону: цей регулятор кодує білок, званий репресором, який, здається, зв’язується з ДНК у гені -операторі. між промотором і структурними генами фактично блокує продукцію мРНК.
Репресор, у свою чергу, управляється, а управління здійснюється за допомогою «сигнальної» речовини. У випадку індукованих ферментів ця речовина є "індуктором. Індуктор" зв'язується з молекулою репресора, змінюючи її форму, так що вона більше не може адаптуватися до ДНК: у цьому випадку, оскільки між промотором і структурними генами немає репресора , репресор може утворювати молекули мРНК і з них молекули білка. З вичерпанням запасу індуктора регулятор знову поверне контроль, що зупинить виробництво нової мРНК, отже, нових білків.У системі бета-галактозидази індуктором є лактоза або речовина, дуже схожа на цю. Похідне: вони приєднається до репресора, інактивуючи його, щоб дозволити біосинтез ферментів. У разі репресивних ферментів речовина, яка діє як "сигнал", діє як корепресор: репресор активний лише у поєднанні з корепресором. У системі гістидину, яка включає в себе десяток різних ферментів, саме ця амінокислота в поєднанні з її тРНК - корепресором - гістидином.
АЛОСТЕРИЧНІ ВЗАЄМОДІЇ
Алостеричні взаємодії, що включають інактивацію ферменту шляхом зміни його форми, забезпечують інший спосіб регулювання метаболічної активності клітини. Алостеричні взаємодії дозволяють здійснювати більш точний контроль, ніж система індуктор -репресор оперону, але не досягають корисного результату виключення біосинтезу даної речовини з першої стадії - продукування мРНК.
СИСТЕМИ УПРАВЛІННЯ В ЄВКАРІЇ
Існують деякі факти, які дозволяють вважати, що система регуляції, подібна до оперону, діє і є вищою серед рослин і тварин. Хромосоми цих організмів сильно відрізняються від хромосом E. coli та інших прокаріотів. Контроль генів у цих Механізм мітозу такий, що кожна клітина даної рослини чи тварини володіє всією інформацією
генетика, присутня в заплідненій яйцеклітині. Тому більшість генів у будь -якій спеціалізованій клітині залишатимуться неефективними протягом усього життя клітини. ДНК у цих клітинах завжди пов'язана з білками. Тому можливо, що репресія генів у еукаріотів вимагає саме цієї асоціації між ДНК і білками.